
Comment réussir un exercice de génétique :
1°) Présenter le croisement
2°) Analyser la génération F1
- De l’homogénéité de cette génération F1 confirmer que les parents sont homozygotes,
- A partir du phénotype de cette génération F1 déduire quels sont les caractères dominants et récessifs,
- Donner les conventions d'écriture (caractère dominant avec une majuscule, récessif avec une minuscule ou caractère sauvage avec « + » ).
3°) Déterminer le nombre de gènes impliqués (s’il n’est pas précisé dans l’énoncé que le caractère est gouverné par un seul gène)
- Analyser le deuxième croisement
- S’il s’agit d’un test-cross analyser les résultats pour chaque caractère :
- si les résultats présentent deux phénotypes alternatifs dans les mêmes proportions on peut en déduire qu’un seul gène est impliqué et qu’il existe deux allèles pour ce gène
- si deux phénotypes alternatifs avec d’autres proportions : plusieurs gènes impliqués
4°) Un seul gène impliqué (monohybridisme)
- Donner les génotypes et les phénotypes des parents et des individus de la F1.
- Effectuer le tableau de croisement du deuxième croisement (F2 ou F’2).
- Comparer les résultats du tableau avec les résultats du croisement donnés dans l'énoncé.
- Conclure
5°) Deux gènes impliqués (dihybridisme) pour un seul caractère ou deux gènes différents pour deux caractère différents : localiser les gènes sur l es chromosomes
- Enoncer les deux hypothèses possibles concernant la localisation des gènes
. H1 les gènes sont liés, ils sont situés sur la même paire de chromosomes.
. H2 les gènes sont indépendants, ils sont situés sur deux paires de chromosomes différentes.
- analyser le deuxième croisement et démontrer qu’il s’agit d’un test-cross (si ça n’a pas été fait précédemment)
- analyser les résultats du test-cross, nommer la génération F’2
- indiquer que le parent double récessif ne produit que des gamètes avec des allèles récessifs
– préciser que les phénotypes de F’2 ainsi que leur fréquence renseignent sur la combinaison d’allèles produits par les individus F1
- si les proportions des combinaisons entre gamètes sont équiprobables : les gènes sont indépendants
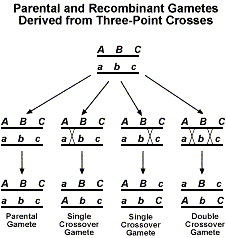
- si les combinaisons parentales sont beaucoup plus fréquentes que les recombinaisons : les gènes sont liés et les recombinants sont dus à des crossing-over au cours de la méiose
- donner les génotypes et les phénotypes des parents et des individus de la F1 en suivant l'hypothèse retenue.
- effectuer le tableau de croisement et expliquer les résultats de ces croisements.
Comparer les résultats du tableau avec les résultats du croisement donnés dans l'énoncé et valider ou rejeter votre hypothèse.
Pour les mécanismes chromosomiques :
- gènes liés : brassa intrachromosomique suivi d’un brassage interchromosomique
- gènes indépendants : brassage interchromosomique
6°) Synthèse : récapitulation rapide afin de fournir une réponse courte au problème posé.
ATTENTION : à adapter selon les exercices.
Dans certains cas on commence l’exercice par un croisement d’individus qui ne sont pas de souches pures, ou par un test-cross …















{kind=link}
0 commentaires :